Inspired by Chris Chambers and
Mark Stokes, this is a research briefing about a recent paper published by
myself and Neil Burgess. My hope is that it explains why we did the research,
what we found, and why we feel this is important. As with the research
briefings from the names above, it is aimed at a scientifically-curious
individual without any prior knowledge in the subject. In relation to this, I
hope I pitch it correctly – do let me know one way or the other!
Horner, A.J., & Burgess, N. (2013) The associative structure of memory for multi-element events, Journal of Experimental Psychology: General, 142(4), 1370-1383. [abstract & PDF]
When we remember previous events
in our lives we bring to mind a wealth of information. We remember the layout
of the living-room, the person we were talking to, the music that was playing
in the background and the smell that wafted through from the kitchen. When we
first experienced this event, each element within this multi-sensory experience
was processed in different regions of the brain. For example, the visual
information will have been processed in visual cortex in the occipital and
temporal lobes and the auditory information in auditory cortex in the temporal
lobes. How is it that we are able to remember all these multiple elements despite
them being processed in different regions of the brain?
Theories of episodic memory
(memory for events) suggest that all the multiple elements of an event are
bound together in an ‘event’ memory (or, more precisely, and event ‘engram’ or event
‘memory trace’) [1]. By binding all this information into a single engram, all the
elements from an event can be retrieved and re-experienced at a later time. The
hippocampi, located in the temporal lobes, are thought to support this binding
as they receive input from multiple regions of the cortex. In this sense, they
act as ‘convergence zones’, binding information across our multiple senses [2].
Although most theories of
episodic memory presume the existence of bound event engrams, little evidence
has been presented in support of this idea [though see 3]. One prediction that stems from this
proposal is that the retrieval of elements from the same event should be
related. If you retrieve information about the location you were in, you should
be more likely to retrieve information about who was there and what they were
saying at the time. In other words, retrieval of an event should be ‘all-or-none’
in nature. We tested this prediction, providing support for the existence of bound
event engrams.
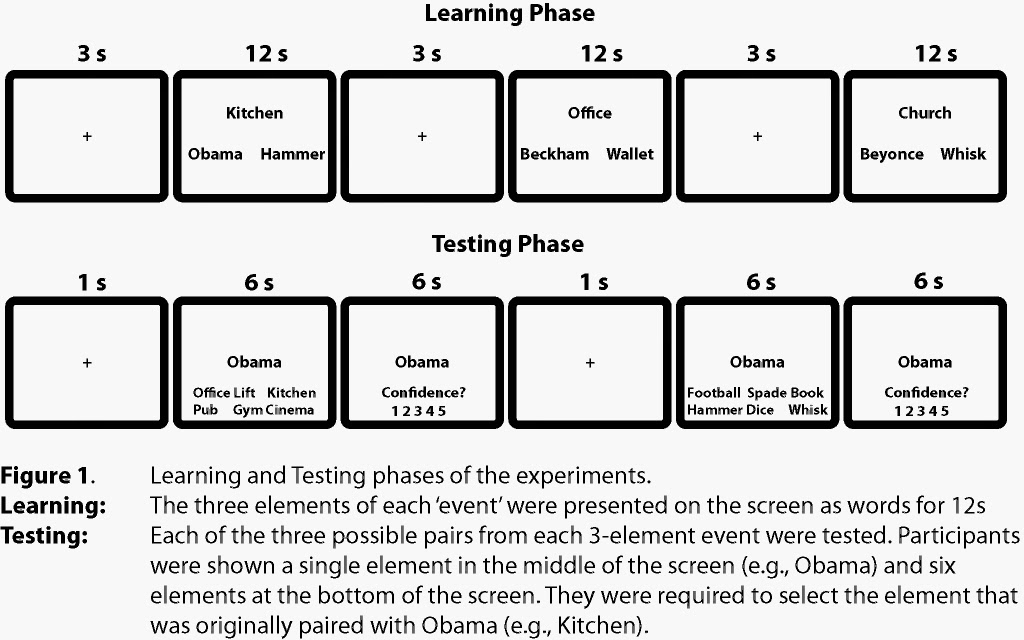
The experiments consisted of a learning
and a testing phase (Figure 1). During learning, participants were presented
with a series of ‘events’. Each event consisted of three elements – a location, famous
person and object. For example, an event might be: Kitchen, Barack Obama and
Hammer. The three elements of each event were presented on a computer screen,
as words. Participants were required to construct a mental image of the three
elements interacting and imagine it as vividly as possible. For example, they
might imagine Barack Obama smashing kitchen cupboards with a hammer. After
learning several events we tested their memory for each pairwise association
within each three element event. In the above example, this meant testing the
association between Kitchen and Barack Obama; Kitchen and Hammer; and Barack
Obama and Hammer.
 If all three elements of an event
are bound in a single event engram, performance for each pair should be related.
If you remember Kitchen and Barack Obama, you should also remember Kitchen and
Hammer. Equally, if you don’t remember one pair, you shouldn’t remember the
other pairs from the same event. We tested this ‘dependency’ between retrievals
from the same event. In order to control for differences in accuracy and the
level of guessing across participants, we developed models that predicted the
level of dependency if performance of pairs for an event were unrelated (the
Independent Model) or if performance of pairs for an event were completely
related (the Dependent Model). We then compared the amount of dependency for
each participant with each of these models to see if the retrieval of elements
from the same event were related. Across three experiments we showed strong
evidence for this predicted dependency. The level of dependency was
consistently greater than the Independent Model (Figure 2), and in some situations did not
differ from the Dependent Model (see paper).
If all three elements of an event
are bound in a single event engram, performance for each pair should be related.
If you remember Kitchen and Barack Obama, you should also remember Kitchen and
Hammer. Equally, if you don’t remember one pair, you shouldn’t remember the
other pairs from the same event. We tested this ‘dependency’ between retrievals
from the same event. In order to control for differences in accuracy and the
level of guessing across participants, we developed models that predicted the
level of dependency if performance of pairs for an event were unrelated (the
Independent Model) or if performance of pairs for an event were completely
related (the Dependent Model). We then compared the amount of dependency for
each participant with each of these models to see if the retrieval of elements
from the same event were related. Across three experiments we showed strong
evidence for this predicted dependency. The level of dependency was
consistently greater than the Independent Model (Figure 2), and in some situations did not
differ from the Dependent Model (see paper).
Thus, we provided evidence that
the multiple elements of an event are bound within a single ‘event’ engram.
Event engrams are thought to be the fundamental building blocks of episodic
memory. They allow us to retrieve information about a single event, without
retrieving information from other similar events. Despite the common assumption
that these event engrams exist, we provide some of the first behavioural
evidence for their existence. Our future work is focussing on revealing the
exact neural mechanism that allows for this event binding, as well as investigating
which regions of the brain support this binding process.
- Tulving, E. (1983). Elements of episodic memory. Oxford: Clarendon Press.
- Damasio, A. R. (1989). The Brain Binds Entities and Events by Multiregional Activation from Convergence Zones. Neural Computation, 1(1), 123–132.
- Jones, G. V. (1976). A fragmentation hypothesis of memory: Cued recall of pictures and of sequential position. Journal of Experimental Psychology: General, 105(3), 277–293.

