Horner, A.J., Bisby, J., Bush, D., Lin, W-J., & Burgess, N. (2015) Evidence for holisitic episodic recollection via hippocampal pattern completion, Nature Communications, 6:7462 doi: 10.1038/ncomms8462
Think back to your last birthday.
Perhaps you were at home, eating good food. Perhaps you were in a pub, drinking
good beer. Perhaps you were in a club, dancing to terrible music.
When we recall events like these
from our past we are able to re-immerse ourselves in the experience, as if we
were there once again. You might remember being in your dining room, eating
birthday cake, whilst your friends sing happy birthday. You might even remember
incidental details, like what you were thinking at the time or the music
playing in the background. How do we remember and re-experience these complex
events?
A long-standing theory,
originally proposed by Marr but developed by many others, suggests that the individual
elements of a complex event are represented in distinct neocortical regions.
For example, the faces of our friends might be represented in visual regions in
the ventral temporal lobe whilst the background music might be represented in
auditory regions in the lateral temporal lobe. These distinct elements are
thought to be bound in a single coherent memory – what Tulving referred to as
an ‘event engram’. It is the hippocampus, receiving input from multiple
neocortical regions (acting as a ‘convergence zone’ in the words of Damasio),
that is thought to form these event engrams when we first experience an event.
What happens when remembering
this event at a later date? Perhaps you meet a friend who attended your
birthday party. This friend acts as a ‘cue’ to retrieve the previous event. Importantly,
with a single cue we are able to retrieve the entire event. In this case, we
see our friend and that enables us to remember the room we were in, our
birthday cake, the background music etc. This retrieval of a complete memory
from a partial cue is known as ‘pattern completion’ and is thought to be a key
function of the hippocampus (and particularly subfield CA3 of the hippocampus).
Following this pattern completion process in the hippocampus, all the retrieved
elements are thought to be ‘reinstated’ in the neocortex. In other words, the
same representations that were active when we first experienced an event become
active at retrieval. It is this hippocampal pattern completion process,
followed by reinstatement of all event elements in the neocortex, that is
thought to underpin ‘recollection’ – our ability to subjectively re-experience
a previous life event.
Despite a wealth of evidence for
the involvement of the human hippocampus in episodic memory, and recollection
in particular, evidence has not been presented for this pattern completion
process in relation to the retrieval of complex events.
Participants learnt pairwise
associations of locations (e.g., kitchen), famous people (e.g., Barack Obama),
objects (e.g., hammer) or animals (e.g., dog). Importantly, each pairwise
association overlapped with other associations, forming complex ‘associative
structures’ (see Figure 1). For example, you might learn ‘Kitchen-Obama’ on one
trial, ‘Obama-hammer’ on a second trial and ‘hammer-kitchen’ on a third trial.
As such, we build relationships between multiple elements across separate
encoding trials. This is an example of a ‘closed-loop’ structure, where each
element is paired with each other element (forming a triangle of associations).
This closed-loop condition is compared to ‘open-loop’ structures, where a chain
of three associations is formed between four elements (see Figure 1).
Importantly, both conditions are formed from three pairwise associations across
three encoding trials. Participants are asked to vividly imagine the two
elements for each association ‘interacting in a meaningful way’.

At retrieval we tested each
pairwise association. For example, we cued with ‘Obama’ and participants were
required to retrieve ‘kitchen’. They were shown six elements of the same type
(locations in this example) and asked to select the element (kitchen)
originally paired with the cue (Obama). The retrieval trials were identical for
both the closed-loop and open-loop condition.
How does this allow us to look
for pattern completion? If pattern completion is present then when retrieving a
single element, all other elements should also be retrieved. In our example,
when cued with ‘Obama’ and retrieving ‘kitchen’ the object associated with
these two elements (‘hammer’) should also be retrieved. This is despite
‘hammer’ being task-irrelevant during this trial.
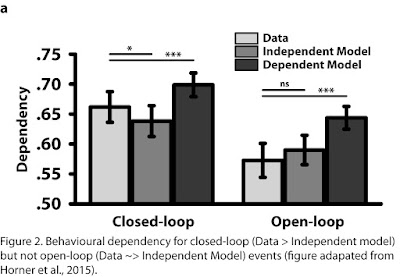 This retrieval should have behavioural
consequences – retrieval accuracy for any two elements within an event should
be related (called ‘behavioural dependency’). If you successfully retrieve
‘kitchen’ when cued with ‘Obama’, you should be more likely to retrieve
‘hammer’ when cued with ‘Obama’. This is because your retrieval success for one
element is based on the strengths of all the associations for a single event. We
provide evidence for this ‘behavioural dependency’ in our closed-loop, but not
open-loop, condition (see Figure 2). This suggests, despite their similarity at
both encoding and retrieval, that pattern completion is present in the
closed-loop but not the open-loop condition.
This retrieval should have behavioural
consequences – retrieval accuracy for any two elements within an event should
be related (called ‘behavioural dependency’). If you successfully retrieve
‘kitchen’ when cued with ‘Obama’, you should be more likely to retrieve
‘hammer’ when cued with ‘Obama’. This is because your retrieval success for one
element is based on the strengths of all the associations for a single event. We
provide evidence for this ‘behavioural dependency’ in our closed-loop, but not
open-loop, condition (see Figure 2). This suggests, despite their similarity at
both encoding and retrieval, that pattern completion is present in the
closed-loop but not the open-loop condition. 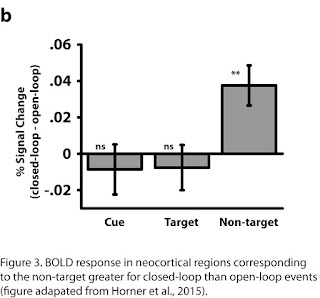 If pattern completion is present
in the closed-loop condition we should see reinstatement of all elements in the
neocortex – including the ‘non-target’ element. Using fMRI, we identified
neocortical regions associated with the encoding/retrieval of individual
elements. Locations were associated with the parahippocampal gyrus, famous
people with the medial prefrontal cortex and objects/animals with lateral
occipital cortex. We next looked for ‘reinstatement’ of non-target elements. If
cuing with a location, and retrieving a person, we should also see
reinstatement in the region associated with objects/animals. We found greater
activity in non-target regions for closed-loops relative to open-loops, again
consistent with pattern completion in the closed-loop but not open-loop
condition (see Figure 3). Importantly, we also show this ‘behavioural
dependency’ and retrieval of non-target element in a computational model of the
hippocampus (an attractor network model), further demonstrating the presence of
pattern completion in the closed-loop but not the open-loop condition.
If pattern completion is present
in the closed-loop condition we should see reinstatement of all elements in the
neocortex – including the ‘non-target’ element. Using fMRI, we identified
neocortical regions associated with the encoding/retrieval of individual
elements. Locations were associated with the parahippocampal gyrus, famous
people with the medial prefrontal cortex and objects/animals with lateral
occipital cortex. We next looked for ‘reinstatement’ of non-target elements. If
cuing with a location, and retrieving a person, we should also see
reinstatement in the region associated with objects/animals. We found greater
activity in non-target regions for closed-loops relative to open-loops, again
consistent with pattern completion in the closed-loop but not open-loop
condition (see Figure 3). Importantly, we also show this ‘behavioural
dependency’ and retrieval of non-target element in a computational model of the
hippocampus (an attractor network model), further demonstrating the presence of
pattern completion in the closed-loop but not the open-loop condition.
Finally, we correlated this
‘non-target’ reinstatement with the BOLD response across the whole brain to see
what other regions correlated with reinstatement. This revealed the hippocampus
(see Figure 4). The BOLD response in the hippocampus correlated (across
participants) with the amount of neocortical reinstatement for the non-target
element. This result supports the idea that the hippocampus is performing
pattern completion, retrieving all event elements, allowing for the
reinstatement of these elements in the neocortex.
What is critical to our study is
that we always compare the closed-loop relative to the open-loop condition. In
both conditions participants have learnt a series of overlapping pairwise
associations and are successfully performing pairwise associative retrieval. As
such, all our results are related to processes over-and-above simple pairwise
associative retrieval. It is this careful experimental design that we believe
is critical to our ability to infer the presence of pattern completion in our
data.
To summarise, we have presented behavioural,
computational modelling and fMRI evidence for hippocampal pattern completion
and neocortical reinstatement in humans, and related these processes to the
retrieval of complex events. We believe this is the first evidence to support a
long-standing mechanistic account of recollection – our ability to subjectively
re-experience previous life events.